Invented by Ricardo Azpiroz, Sunghwa Choe, Kenneth A. Feldmann, Arizona Board of Regents of University of Arizona
In recent years, there has been a growing interest in the field of biotechnology, particularly in the development and application of polynucleotides and polypeptides. One such class of molecules that has gained significant attention is DWF4 polynucleotides and polypeptides. These molecules have shown immense potential in various industries, including agriculture, pharmaceuticals, and biofuels.
DWF4, or Dwarf4, is a gene that plays a crucial role in regulating plant growth and development. It is responsible for the biosynthesis of brassinosteroids, a group of plant hormones that control cell elongation and division. By manipulating the expression of the DWF4 gene, scientists have been able to develop polynucleotides and polypeptides that can modulate plant growth, leading to improved crop yields and enhanced stress tolerance.
One of the most significant applications of DWF4 polynucleotides and polypeptides is in agriculture. With the global population expected to reach 9.7 billion by 2050, there is an urgent need to increase food production. By using DWF4-based technologies, farmers can cultivate crops that are more resistant to environmental stresses such as drought, heat, and salinity. These technologies can also lead to the development of dwarf varieties of crops, which require less space and resources, making them ideal for urban farming and vertical agriculture.
The pharmaceutical industry is another sector that can benefit from the use of DWF4 polynucleotides and polypeptides. The ability to modulate plant growth and development can be harnessed to produce high-value secondary metabolites, such as pharmaceutical compounds. By manipulating the expression of the DWF4 gene, scientists can enhance the production of these compounds, leading to the development of new drugs and therapies. Additionally, DWF4-based technologies can be used to produce plant-based vaccines, which offer a safer and more cost-effective alternative to traditional vaccine production methods.
Furthermore, DWF4 polynucleotides and polypeptides have the potential to revolutionize the biofuel industry. With the increasing concerns about climate change and the need to reduce greenhouse gas emissions, there is a growing demand for sustainable and renewable energy sources. By manipulating the expression of the DWF4 gene, scientists can enhance the production of biomass in plants, which can be used as a feedstock for biofuel production. This can lead to the development of more efficient and environmentally friendly biofuels, reducing our dependence on fossil fuels.
In terms of market size, the global market for DWF4 polynucleotides and polypeptides is expected to witness significant growth in the coming years. The increasing demand for high-yielding and stress-tolerant crops, as well as the need for innovative pharmaceutical and biofuel solutions, are driving the market. Additionally, advancements in genetic engineering techniques and the increasing adoption of biotechnology in agriculture and pharmaceutical industries are further fueling the market growth.
However, it is important to note that the commercialization of DWF4 polynucleotides and polypeptides is still in its early stages. There are several regulatory and ethical challenges associated with the use of genetically modified organisms, which need to be addressed before widespread adoption. Additionally, the high costs associated with research and development, as well as the limited awareness among end-users, may hinder the market growth to some extent.
In conclusion, the market for DWF4 polynucleotides and polypeptides is poised for significant growth in the coming years. The ability to modulate plant growth and development using these molecules offers immense potential in agriculture, pharmaceuticals, and biofuels. However, it is crucial to address the regulatory and ethical challenges and increase awareness among end-users to fully realize the benefits of these technologies. With continued advancements in biotechnology, DWF4-based solutions have the potential to revolutionize various industries and contribute to a more sustainable future.
The Arizona Board of Regents of University of Arizona invention works as follows
The present invention is a novel polynucleotide isolated from dwarf plants. The dwf4 oligonucleotides encode all or a part of a DWF4 peptide. This is a cytochrome P450, an enzyme that mediates several steps in the synthesis of brassinosteriods. The present invention also pertains to isolated polynucleotides encoding regulatory regions of Dwf4. “Uses of the polypeptides or polynucleotides encoding dwf4 are also disclosed.
Background for DWF4 Polynucleotides and polypeptides, and their Uses
Plant growth is achieved by tightly controlled cell division. Cell expansion is more important in plants than it is for most other organisms. All plan organs are the result of a period where cells elongate, usually following active cell division. The sessile nature requires plants to make rapid but fine adjustments to their growth in order to adapt to harsh environmental conditions. Plasticity in plants, D. H. Jennings & A. J. Trewavas (Cambridge UK: Company of Biologists Ltd), pp. 31-76).
In Arabidopsis cell elongation plays a major role in the growth of hypocotyls in germination seedlings, and in the extension of inflorescences at vegetative growth’s end (bolting). Both external stimuli as well as internal mechanisms are responsible for the coordinated control of plant growth. The most obvious external signal is light (Deng X.W. (1994), Cell 76:423-426). “Light inhibits hypocotyls and promotes cotyledon growth and leaf development, and photoperiod plays a crucial role in flower initiation for many species.
The internal components of signaling in plants are generally mediated through chemical growth regulators. (Phytohormones, reviewed by Klee, H., and Estelle, M., (1991), Annu. Rev. Plant Physiol. Plant Mol. Biol. 42:529-551). Evans,?Functions and organization of hormones in the cell? Hormonal Regulations of Plant Physiology by T. K. Scott (Berlin, Springer-Verlag), pp. 23-79). Growth depends on regulated cell events such as divisions, elongations, and differentiation.
Gibberellic acids (GA) and Cytokinins Promote Flowering. GA also stimulates stem lengthening, while cytokinins reduce apical dominant by increasing axillary shot formation. Auxins, on the other hand, promote apical dominant growth and encourage elongation. This is a result of a process that requires acidification of cell walls by a H+-pumping ATPase dependent on K (Rayle, D. L. and Cleland (1977) Curr. Top. Dev. Biol. 11:187-214).
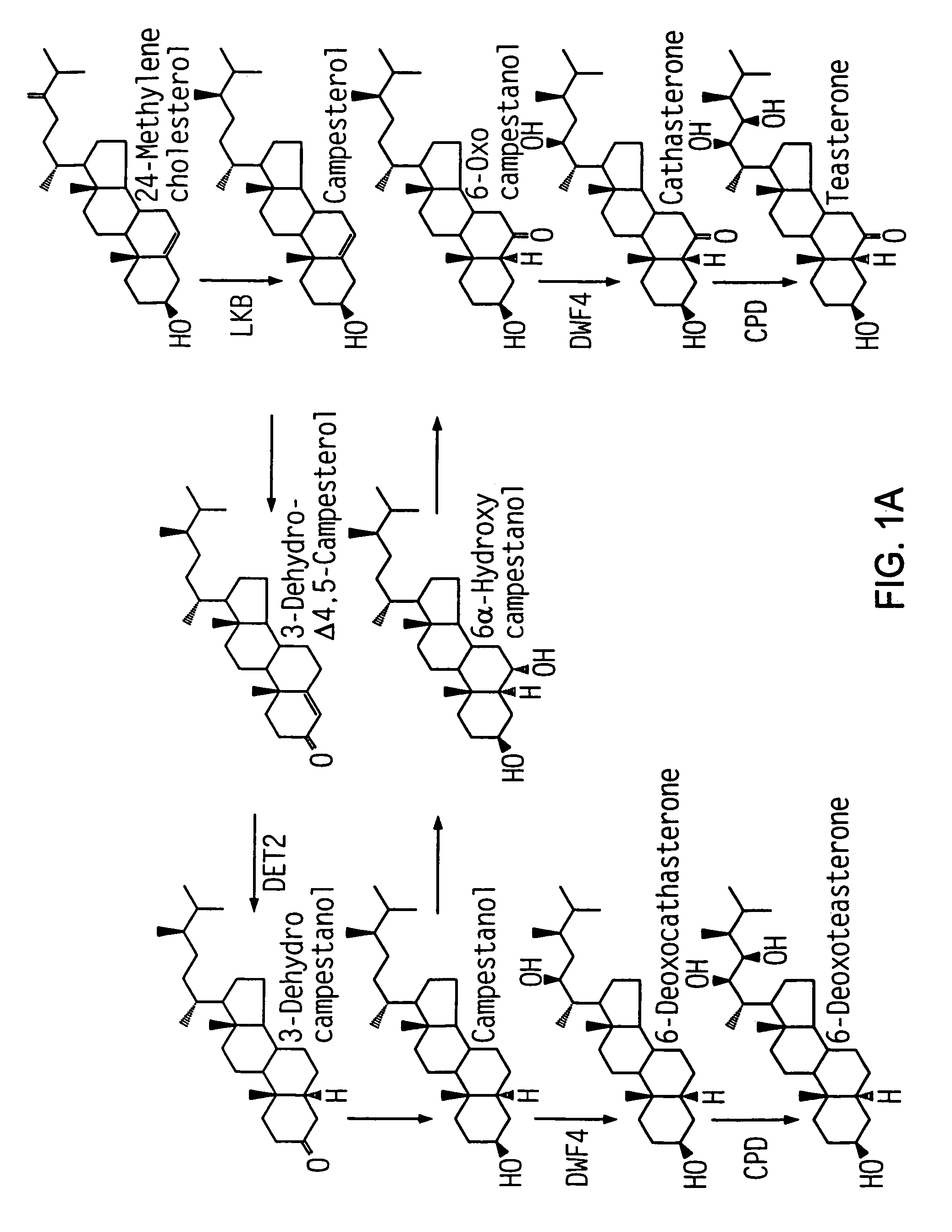
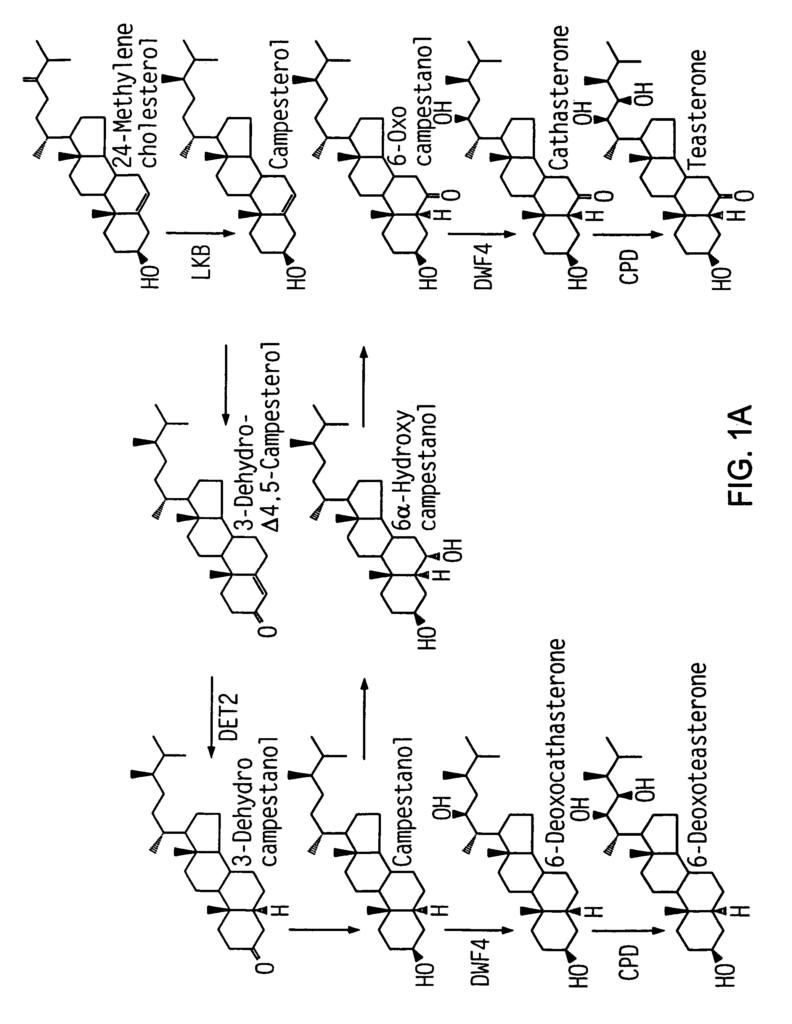
Clouse (1996), Plant J. 10:1-8. 10:1-8). The BRs are the most recent class of plant growth substance discovered. They have been the least studied to date, but rapid progress is being made in understanding BR biosynthesis, regulation, and the like (Yokota T. (1997) Trends Plant Sci. 2:137-143). The term BRs refers to all the growth-promoting steroid found in plants. (Grove et.al. Nature 281, 216-217 (1979). The molting hormones in insects are ecdysteroids. (Richter & Koolman, 1991) “Antiecdysteroid Effects of Brassinosteroids on Insects?” Brassinosteroids, Chemistry, Bioactivity, And Applications, H. G. Cutler, T. Yokota, G. Adam, eds. (Washington, D.C., American Chemical Society), pp. Active BRs, however, have unique structural characteristics. FIG. As shown in FIG. Hydrogen, multiple hydroxylations in four different positions and stereochemistry specific to each position have been suggested as essential configurations for BRs. (Reviewed in Marquardt & Adam (1991),?Recent Advances in Brassinosteroid Research? Chemistry of Plant Protections (W. Ebing, ed., Berlin: Springer-Verlag), pp. 103-139). Among>40 naturally occurring BRs, brassinolide (BL; 2?, 3?, 22(R), 23(R)-tetrahydroxy-24(S)-methyl-B-homo-7-oxa-5?-cholestan-6-one) has been shown to be the most biologically active (reviewed in Mandava (1988) Annu. Rev. Plant Physiol. Plant Mol. Biol. 39:23-52). As a major effect on the body, BRs promote longitudinal growth in young tissues by cell elongation, cell division, and cell division. (Reviewed in Clouse, 1996, supra; Fujioka, and Sakurai, 1997a, Nat. Prod. Rep. 14:10).
Although there is less understanding of the brassinosteriod (BRs Mitchell et al. (1970) Nature 225:1065-1066; Grove et al. Nature 281, 221: 216-217 (1979); Mandava, N. (1988) Annu. Rev. Plant Physiol. Plant Mol. Biol. Biol. Fujioka, S. and Sakurai A. (1997) Physiol. Plant 100:710-715).
Arabidopsis has been reported to have several types of dwarf-like mutants. There are a number of mutations that have been found to affect hormone (axr2) or light-dependent pathways (cop, det and fusca, another mutant group with members who show perturbations in light-regulated cell growth). These mutations also cause defects in the elongation of cells. These mutants are also characterized by other phenotypic changes. At least five GA mutations have been described to be smaller (Koornneef & Van der Veen, 1980) Theor. Appl Genet. 58:257-263).
Another hormonal mutation, auxin-resistant2 (axr2) results in plants that are dwarf in both the light and dark as well as having increased resistance to high concentrations of auxin and ethylene. Planta 188, 271-278 (1992). There is an interesting relationship between light regulation levels and cytokinins. Arabidopsis seeds grown in darkness in the presence cytokinins open their cotyledons and initiate leaf development. They also activate transcription of the chlorophyll-a/b binding proteins gene (CAB). They also show a cytokinin-dependent dwarf phenotype.
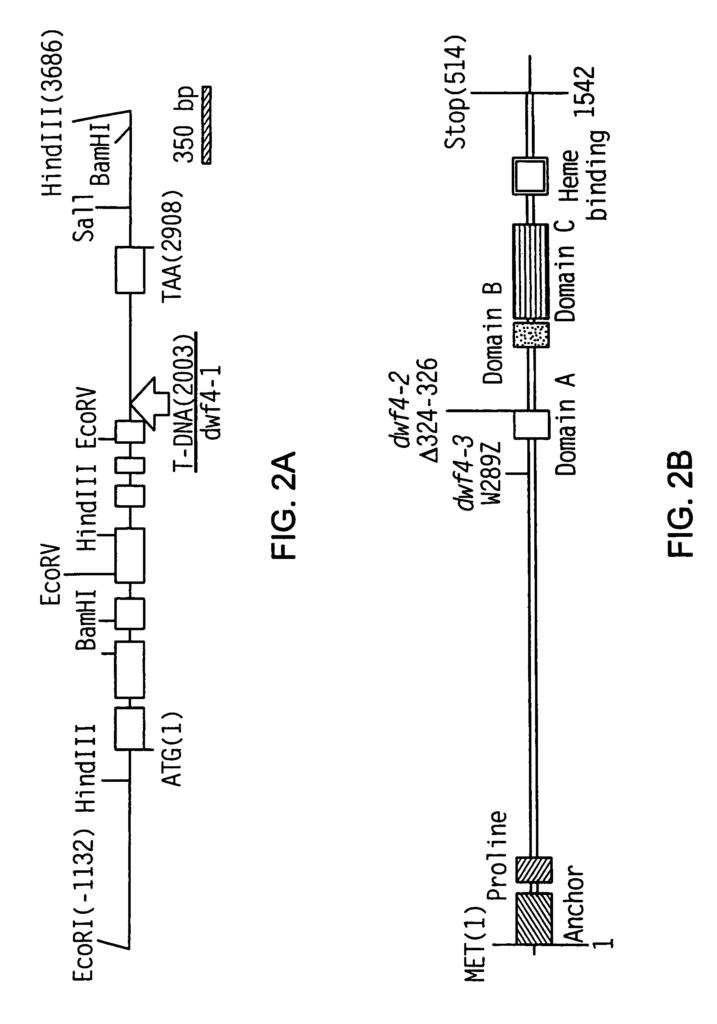
Kauschmann et. al. have described dwarf Arabidopsis mutations that can be rescued with the addition of BRs. (1996) Plant J 9:701-713; Li et al. (1996) Science 272:398-401; Szekeres et al. (1996) Cell 85:171-182; Azpiroz et al. Plant Cell 10 (1998) 219-230, including three mutants dwarf1 (dwf1 Kauschmann and al. (1996) Plant J. Plant J. 9:701-713), constitutive morphogenesis (cpd, Szekeres and al. Cell 85:171-182 (1996) Szekeres et al. 91996) Science 272 (p. 398-401). These mutants are defective in steroid synthesis. DWF1 (Feldmann et al. First, DWF1 (Feldmann et al. (1989), Science 243:1351-1354), was cloned (GenBank accession U12400). Takahashi et al. (1995) Genes Dev. They hypothesized in 9:97-107 that DWF1, which was isolated using an allele of dwf1, referred as diminuto1(dim 1), contained a nuclear targeting signal which could confer a regulation function to the protein. Mushegian & Koonin (1995), Protein Sci. According to 4:1243-1244, DWF1 shows limited homology with the flavin adenine dinucleotide independent oxidoreductase (FAD), suggesting that DWF1 has an enzymatic role in BR biosynthesis. Kauschmann and colleagues report that. 1996), supra (dwf1-6 were described as cabbage1 (cbb1)), dwf1 mutations were saved by the exogenous addition of BRs.
DET2 encodes a putative steroid 5-reductase that mediates an early step in BR Biosynthesis (Li et. (1996) supra, Li et al. (1997) Proc. Natl. Acad. Sci. USA 94:3554-3559; Fujioka et al. Plant Cell 9 (1951-1962) (1997); FIG. 1). Moreover, det1-det2 are less dependent on cytokinins and seem to be saturated in a cytokinindependent delay of senescence in tissue culture (Chory et al.). 91994) Plant Physiol. 104:339-347). CPD is a novel cytochrome P450, (CYP90A1 – Szekeres et. al. (1996), supra), encoding a possible 23?-hydroxylase which acts in BR Biosynthesis. The deetiolated mutants (det) as well as the constitutive photomorphogenic mutants (cop) are phenotypically diverse. Mutations of DET1, COP1, COP8, COP9, COP10 and COP11 lead to constitutive derepression of large portions of the photomorphogenic programme (Chory et. al. Plant J. 1992; Deng X-W. and quail P. H. Wei, N. and Deng, X.W. (1992) Plant Cell 4:1507-1518; Wei et.al. Wei et al. Plant Cell 5 (93), 329-339. This class of mutants that regulate light is the only one to show a significant reduction in height both in light and in darkness.
There are other dwarfs who are insensitive (bri) to one of these hormonal compounds. (1996) Plant Physiol. Li and Chory, Cell 90:929-938 (1997)), gai (gibberellic-acid insensitive. Koornneef. (1985) Physiol. Plant. Plant. Genetics 138, 1239-1249 (1994). Clouse et al. (1996), supra isolated bri by screening ethyl methanesulfonate-mutagenized populations for mutants whose root growth is not retarded at inhibitory concentrations of BR. The BR1 is implicated in BR signal transduction or perception (Clouse, 1996, supra). Kauschmann et al. (1996), supra describe a mutant cbb2 with phenotypically identical phenotype that maps at the same location. The dwf2 variants also have a similar phenotype to bri, and map in the same location (Feldmann & Azpiroz 1994) “dwarf” (dwf), and “twisted dwarf” (twd). In Arabidopsis, An Atlas of Morphology and Development (J Bowman, Ed., New York: Springer-Verlag), pages 82-85. 82-85). All of the BR insensitive dwarf mutants that have been described so far are likely allelic. Recent BRI cloning and analysis has shown that it encodes a leucine rich-repeat receptor kinase. This suggests a role for the BR signaling pathway.
Other plant species have also been found to contain mutants that are defective in BR Biosynthesis. Bishop et al. Plant Cell 8:959-969 (1996) isolated a tomato mutant using transposon tag. The tomato Dwarf encodes a pioneer member of the CYP85 Family, and appears to be involved with BR biosynthesis. In addition, Nomura et al. (1997) Plant Physiol. 113:31-37 stated that “the lka and lkb mutations in garden pea were deficient in BR perception (lka) or biosynthesis (lkb).
At the moment, we know very little about the downstream processes that take place in response to these signalling events and control directly cell size. It is difficult to understand the biochemical processes and cell biology involved. There is also little information on the coordination of the different signaling pathways that are convergent at the cell and how they are coordinated. It is unclear how hormones and light interact to control cell elongation. There is still a need to identify and characterize additional mutants, and polypeptides that are encoded by these mutants in the pathways of plant growth.
In one aspect, the invention comprises an isolated dwf4 oligonucleotide that has an open reading frame and encodes a peptide containing (i) sequences that are more than 43% identical to 10 consecutive amino acids in SEQUENCE NO:2 or (ii), sequences that contain at least 10 contiguous residues that have a greater than 44 % identity to SEQUENCE NO:2, a complement or reverse compliment of said polynucleotide. In some embodiments, the polynucleotide has at least 70% similarity to the DWF4 coding region in SEQ ID No:1 and its complements or reverse complements. In other embodiments, the isolated polynucleotide dwf4 contains the nucleotide of SEQ NO:1, its complements and reverse compliments. The polynucleotide can also contain at least 30 consecutive SEQ ID No:1 nucleotides.
In another aspect of the invention, an isolated dwf4 nucleotide is included. This polynucleotide consists (i) of a sequence that shares at least 50 percent identity with SEQID NO:1, its complements and inverse complements or (ii), a sequence containing at least 15 contiguous dwf4 nucleotides and sharing at least 50 percent identity with SEQID NO:1, its complements and inverse complements. In some embodiments, the isolated dw4 has a minimum of 50% identity with the DWF4 coding region in SEQ ID No:1, its complements and reverse counterparts. In other embodiments, isolated dwf4 oligonucleotides comprise nucleotide sequencing of SEQ NO:1, its complements and reverse counterparts or nucleotide segments containing at least 30 consecutive SEQ NO:1 nucleotides. The dwf4 DNA polynucleotides may be genomic and include introns. In other embodiments, a dwf4-control element is included in the polynucleotide. This element comprises a polynucleotide chosen from the group consisting (i), a sequence with at least 50% similarity to the nucleotides 1-3202 of SEQID NO:1 (ii), a fragment (i), which contains a control element (iii), and (iii), complements or reverse complements (iii). In yet further embodiments, a dwf4 element is included in the polynucleotide. This includes a polynucleotide that has at least 50% identity with nucleotides corresponding to 3? In still further embodiments, the polynucleotide includes a dwf4 control element comprising a polynucleotide selected from the group consisting of (i), a sequence having at least 50% identity to nucleotides 6111 to 6468 corresponding to SEQ ID No:1’s 3? UTR, and (iii), complements or reverse complements of either (i) (iii). In certain embodiments, a polynucleotide comprises a dwf4 oligonucleotide that is selected from a group consisting (i), a sequence with at least 50% similarity to the sequences which correspond to the introns in SEQID NO:1 (ii), a fragment (i), which includes a Dwf4 intro (iii), and (iii), complements and reverse compliments of the (i), (ii 54. “In still further embodiments any of the polynucleotides herein described can be operably linked to nucleic acids encoding heterologous polypeptides (e.g. a polypeptide cytochrome P450), for example as a hybrid polynucleotide.
In another aspect of the invention, recombinant vectiosn are recombinant polynucleotides that contain a dwf4 control element described above (e.g., promoter or introns) and control elements linked to them, allowing a coding segment within the polynucleotide to be transcribed into a host cell. In certain embodiments the recombinant molecule consists of (a) polynucleotides that include the dwf4 (e.g. promoters or introns) control element. (b) A nucleic acids molecule containing a coding element operably linked to the control element.
Host Cells comprising or transformed with any of recombinant vec- tors described herein” are also provided. In some embodiments the host cells are grown ex vivo, while in others the dwf4 nucleotide can be provided to the host cell intra vivo. In certain embodiments, the DWF4 is supplied in quantities that allow a plant to regenerate.
In another aspect of the invention, it includes a modulation method for a DWF4 peptide that comprises the following steps: a) providing a cell host as described in this document; and b) culturing the cell host under conditions such that the dwf4 oligonucleotide contained within the cell host is transcribed. In some embodiments, dwf4 is expressed in excess. In other embodiments, polynucleotides included in host cells inhibit expression of dwf4.
In a further aspect, the invention also includes a transgenic crop containing any of the described recombinant vec- tinants.
In another aspect of the invention, a method for producing a polypeptide recombinant is provided. This includes the steps: (a), providing a host-cell as described in this document; and (b), culturing the host-cell under conditions that allow the expression of the recombinant peptide encoded by coding sequences present in the recombinant vector.

Click here to view the patent on Google Patents.
Leave a Reply