Invented by Casey C. Case, George N. Cox, III, Stephen P. Eisenberg, Qiang Liu, Edward J. Rebar, Sangamo Therapeutics Inc
The Sangamo Therapeutics Inc invention works as follows
The present invention provides methods to modulate expression of endogenous cell genes using engineered Zinc Finger Proteins.

Background for Regulation of endogenous gene expression in cells using zinc finger proteins
The selective up- or downregulation of gene expression can control many, if not most physiological and pathophysiological functions. Pathologies would be treatable if there were methods to control gene expression. Some examples include inappropriate expressions of pro-inflammatory cytokines, such as in rheumatoid arthritis, or underexpression of the hepatic LDL in hypercholesteremia. Other examples are the overexpression of proangiogenic and antiangiogenic growth factors, in solid tumors. Altering gene expression could also be used to control pathogenic organisms, such as bacteria, viruses, fungi and protozoa. “There is a need for simple therapeutic approaches which can up-regulate good genes and down-regulate the disease-causing genes.
This ability can also be used to experimentally determine the function of an interest gene. This is in addition to the direct therapeutic value provided by the ability of manipulating gene expression. In order to determine the function of newly discovered genes, it is common practice to clone their cDNA in an expression vector with a strong promoter. Then the cell that has been transfected will be measured for the physiological effects of the overexpression. This method is labor-intensive and does not consider the physiological effects of down-regulating a target gene. The scientific community would benefit greatly from simple methods that allow the selective over- and underexpression of uncharacterized gene. The scientific community would benefit greatly from simple methods that allow the regulation of genes within cell model systems, transgenic organisms and transgenic plants.
Another use of tools that allow the manipulation of gene-expression is the production of commercially valuable biological products. Transgenic animals, plants and cell lines could be engineered in order to over-express useful protein products. An engineered cell-line can produce erythropoietin. The production of metabolic pathways could also be improved or altered by selectively up- or down-regulating a gene that encodes a key enzyme. This can be achieved by producing plants with different levels of fatty acids saturation.
Methods exist that allow you to change the expression of any gene. For example, by using ribozymes or antisense technology. The methods used to date have been found to be insufficient in many cases and have typically not shown high efficacy in terms of target or specificity. These characteristics are desirable for useful experimental results as well as therapeutic treatments.
Gene expression is controlled by alterations to the function of DNA-binding proteins that are sequence specific. These transcription factors. They bind to the gene’s transcriptional initiation point, usually in close proximity but sometimes at a distance. They influence the formation or function efficiency of a transcription initiating complex at the promoter. Transcription factors may act in either a positive (transactivation), or a negative (transrepression) fashion.
Transcription Factor Function can be constitutive or conditional (always on? “Transcription factor function can be constitutive (always?on?) The conditional function of a transcriptional factor can be conferred by various means. However, the majority of these regulatory mechanism depend on the sequestering the factor in cytoplasm, and its subsequent inducible release, nuclear translocation and DNA binding, and transactivation. Progesterone receptors and sterol-response element binding proteins (SREBPs), as well as NFkappa B, are examples of transcription factors which function in this manner. Some transcription factors respond to phosphorylation by changing their ability to bind the DNA recognition sequence. (Hou et. al. Science 256:1701, 1994; Gossen & Bujard PNAS 89, 5547, 1992; Oligino et. al. Gene Ther. 5:491-496 (1998); Wang et al., Gene Ther. 4:432-441 (1997); Neering et al., Blood 88:1147-1155 (1996); and Rendahl et al., Nat. Biotechnol. 16:757-761 (1998)). This mechanism is found in both prokaryotes and eukaryotes.
Zinc finger proteins (?ZFPs?) “Zinc finger proteins (?ZFPs?)” are proteins that bind DNA in a specific sequence. The transcription factor TFIIIA was first discovered in the oocytes from the African clawed Toad (Xenopuslaevis). ZFPs can be found in many eukaryotic cell types. An exemplary motif characterizing one class of these proteins (C2H2 class) is -Cys-(X)2-4-Cys-(X)12-His-(X)3-5-His (where X is any amino acid). One finger domain has a length of about 30 amino acid and structural studies have shown that the alpha helix contains two invariant residues of histidine, which are coordinated through zinc by the two cysteines in a single beta-turn. Over 10,000 zinc finger sequences were identified to date in over a thousand transcription factors that are known or presumed. ZFPs play a role in DNA recognition, RNA binding, and protein-protein interactions. According to current estimates, this class of molecules is expected to make up about 2% of human genes.
The X-ray structure of Zif268, which is a three finger domain of a murine transcriptional factor, was solved with its cognate sequence of DNA. It shows that the fingers can be superimposed by rotating and translating the finger in a regular pattern along the DNA main axis. The structure indicates that each finger interacts with DNA independently over three base-pairs. Side-chains located at positions?1,?2, 3, and?6 on each recognition-helix make contact with the respective DNA triplet-subsite. The amino terminus is located at the 3? The amino terminus of Zif268 is located at the 3? Recent studies have shown that zinc fingers can bind a fourth base within a target segment. (Isalan, et. al. PNAS 94:5617?5621 (1997). The fourth base is located on the opposite strand to the three other bases that zinc finger recognizes and is complementary to the base 3? The three subsites of the base.
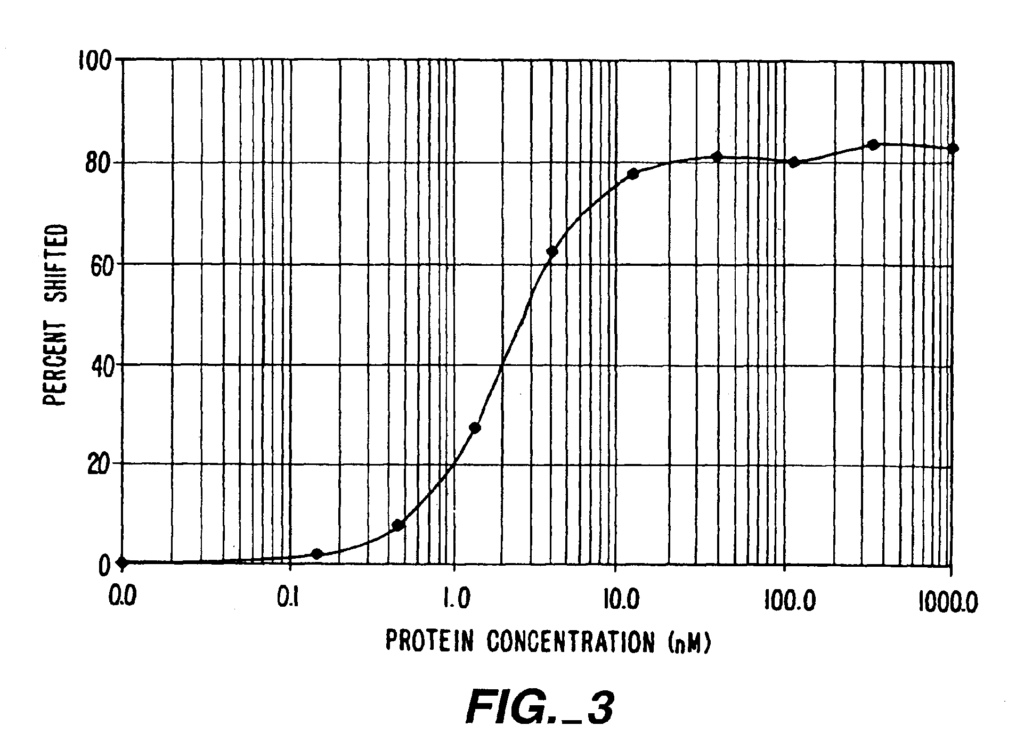
The structure of the Zif268 DNA complex suggested that the DNA sequence-specificity of a ZFP could be altered by substituting amino acids at the four helix position (?1, 2, 3, and 6) of a zinc finger recognition Helix. In a series in 1994, phage display experiments were performed using zinc finger combinatorial library to test this observation (Rebar, et. al. Science 263, 671, 673 (1994); Jamieson, et. al. Biochemistry 33, 5689-5695, (1994); Choo, et. al. PNAS 91, 11163-11167, (1994). Combinatorial Libraries were constructed using randomized side chains in the first or middle fingers of Zif268. The binding sites for Zif268 were then altered by replacing the DNA sub-site with an altered triplet. The correlation between the type of mutations introduced and the alteration in binding particularity led to a partial substitution set for the rational design of ZFPs that have altered binding specificity.
The article “Greisman & Pabo” Science 275:657-661 (2007) discusses an elaboration on a phage-display method where each finger of a Zinc Finger Protein is subjected to successive randomization and selection. This paper reported the selection of ZFPs to target a nuclear hormone receptor element, a p53-target site and a Tata box sequence.

Recombinant ZFPs were reported to be able to regulate the gene expression of transiently-expressed reporter genes in cultures cells (see, for example, Pomerantz and colleagues, Science 267, 95-96; Liu et. al. PNAS 94, 5525-5530, 1997; and Beerli, et. al. PNAS 95, 14628-14633, 1998 )).
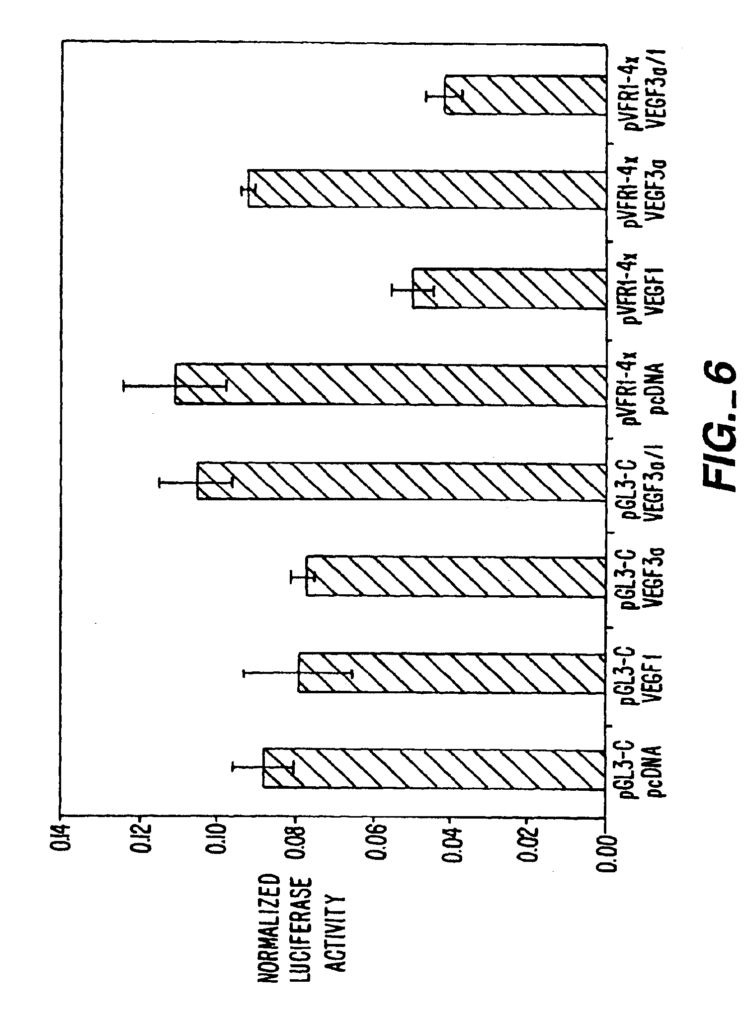
Pomerantz, et. al., Science, 267:93-96, (1995), report on an attempt to create a novel DNA-binding protein by fusing the homeodomain of Oct-1 with two fingers from Zif268. The hybrid protein is then fused to either a transcriptional activator domain or a transcriptional repressor for expression as a “chimeric” protein. The chimeric proteins was found to bind an hybrid target site that represents a subsite of both components. The authors constructed a reporter containing the luciferase operably linked with a promoter, and a hybrid DNA binding protein site in proximity to that promoter. The authors reported their chimeric protein binding DNA could activate or suppress expression of the gene luciferase.
Liu et. al., PNAS, 94:5525-5530, (1997) describe the formation of a composite ZFP using a peptide to link two component ZFPs. Each ZFP has three fingers. The composite protein is then linked to transcriptional activation domains or repression. The chimeric protein formed by the two ZFPs bound target segments was reported to bind to the target site. The chimeric ZFP was also reported to be able to activate or repress transcription when its target site is inserted in a reporter plasmid adjacent to a promoter operably associated with the reporter.
Beerli et. al., PNAS95:14628-14633 (2000) report the construction of a chimeric 6-finger ZFP fused with either a transcriptional repressor KRAB or ERD or SID, or a transcriptional activation VP16 or VP64 domain. This chimeric ZFP is designed to recognize a 18 bp site in the 5′ untranslated region of the human erbB-2 gene. This chimeric ZFP was designed to recognize an 18 bp target site in the 5? The authors of this paper report activation and suppression of a reporter luciferase linked to the erbB-2 gene promoter using this construct.
In addition, a recombinant ZFP has been reported to repress the expression of a plasmid construct encoding a bcr/abl oncogene” (Choo et al. Nature 372:642-645, 1994). The ZFPs were bound to a nine-base sequence GCA GAA GCC that was chosen to overlap a junction created by an oncogenic translocation fusing genes encoding both bcr abl. It was intended that a ZFP binds to the oncogene but not to the abl or bcr components genes. The authors selected a variant ZFP bound to this segment using phage display. This variant ZFP was then reported as repressing expression of a transfected bcr/abl construct.
These methods are primarily focused on the regulation of transiently expressed or exogenous genes integrated into genomes. These transiently expressed gene described by Pomerantz, Liu, and Beerli are episomal genes. The episomal genes are not packaged in chromatin the same way as chromosomal ones. Even the gene described by Choo and colleagues, which is stably expressed, does not fit this description. The random integration of the gene into the genome is different from an endogenous one. “In contrast, the specific regulation of a cellular endogenous gene in its native environment using a ZFP is not demonstrated in the literature.
The present invention provides methods for the first-time of controlling expression of endogenous cell genes in their native chromatin environments, as opposed to genes that are transiently expressed or have been integrated exogenously into the genome.
In one aspect, this invention provides methods to modulate expression of an endogenous molecule in a cell. Modification can be either activation of gene expression or repression. Endogenous genes can be either a cellular, microbial, or viral gene.
The invention provides methods and compositions that modulate expression of an endogenous protein in a cell. This method includes administering a nucleic molecule containing a polynucleotide coding for an engineered Zinc Finger Protein, which comprises a plurality zinc fingers, and wherein at least one zinc finger is derived from information contained in a database containing existing zinc finger proteins with their respective binding affinity.
The inventors have disclosed additional embodiments of their inventions that include methods and compositions of modulating the expression of an endogenous protein in a cell. This method includes administering to the cells a nucleic molecule containing a polynucleotide coding a engineered protein with a plurality zinc fingers, wherein this engineered protein has a plurality zinc fingers, and wherein each of the zinc fingertips is derived from a previously existing zinc finger proteins; and then maintaining the

In further embodiments disclosed herein are compositions and methods for modulating the expression of an endogenous protein in a cell. The method involves administering to the cells a nucleic molecule containing a polynucleotide encoding a zinc finger, wherein said zinc finger comprises a number of zinc fingers, and wherein said zinc finger is partially randomized; and maintaining conditions in the cell in which the engineered protein binds and is expressed at a target site within the gene.